项目文章 | IF=8.4&转录因子Egr-1是脑膜炎型大肠杆菌引起的血脑屏障损伤的关键调节因子

2024年1月17日华中农业大学动科动医学院陈焕春院士、王湘如教授团队在期刊Cell Communication and Signaling(IF:8.4)发表了题为“Egr-1 is?a?key regulator of?the?blood-brainbarrier damage induced by?meningitic?Escherichia coli”的研究论文。该研究结果表明Egr-1依赖性机制是脑膜炎型大肠杆菌破坏血脑屏障的途径,为细菌性脑膜炎的治疗提供了有希望的靶点。爱基百客为该研究提供ChIP-seq技术支持。

? 研究背景??
细菌性脑膜炎仍然是全球感染相关死亡的主要原因。尽管大肠杆菌(E. coli)是新生儿脑膜炎最常见的病因,但感染过程中细菌血脑屏障(BBB)破坏的潜在机制仍然未知。研究者观察到,使用脑膜炎性大肠杆菌感染人脑微血管内皮细胞会触发早期生长反应因子-1(Egr-1)的激活,进而导致血脑屏障的损伤。本文旨在探究Egr-1在脑膜炎型大肠杆菌引发的血脑屏障损伤中的作用和机制。
? 技术路线?

? 研究结果??
1. 细菌感染期间脑内皮细胞 Egr-1 的诱导
为了研究脑膜炎病原体诱导的转录因子模式,研究者对人脑微血管内皮细胞(hBMEC)进行了3小时的感染,结果发现热休克因子(HSF)、肌原调节因子(MYOD)、母体效应蛋白(HEN)、Octamer结合蛋白1(OCT1)和早期生长反应蛋白1(Egr-1)的活性显著增加。在hBMEC感染脑膜炎病原体后,Egr-1的mRNA转录和蛋白水平显著增加。此外,在感染小鼠脑微血管中,Egr-1的转录水平显著上调,蛋白水平也显著增加。这些结果表明,脑膜炎病原体大肠杆菌感染可以显著激活hBMEC中的Egr-1(Fig 1)。
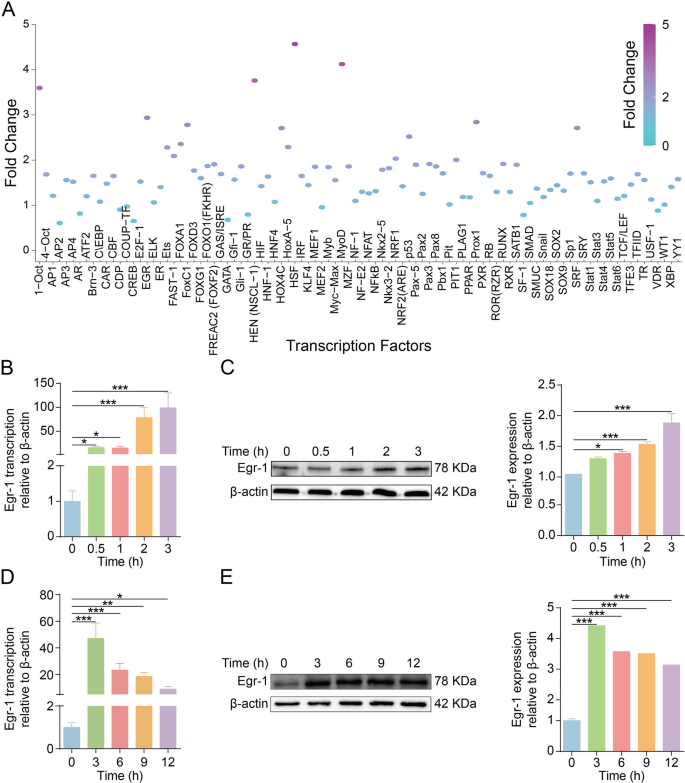
Fig 1.脑膜炎型大肠杆菌感染可显著增加体外和体内Egr-1的激活
2. Egr-1 结合基因的全景图
研究者通过ChIP-seq实验发现在脑膜炎型大肠杆菌感染的人脑微血管内皮细胞(hBMEC)中,Egr-1在细菌诱导的基因组开放位点有明显的富集,表明这些位点受Egr-1调控。统计分析显示,与对照组相比,细菌感染的hBMEC中的可接近峰数量显著增加。在这些peak峰中,感染组和对照组分别有2,562个和99个独特的峰,同时还有137个重叠的峰。进一步利用HOMER分析可接近区域的基因组功能元件,结果显示,感染组中超过80%的峰主要富集在启动子区域,而对照组中的大多数峰富集在远端间基因区域。这些发现表明,Egr-1是脑膜炎型大肠杆菌诱导的血脑屏障功能障碍的主要转录调节因子(Fig 2)。
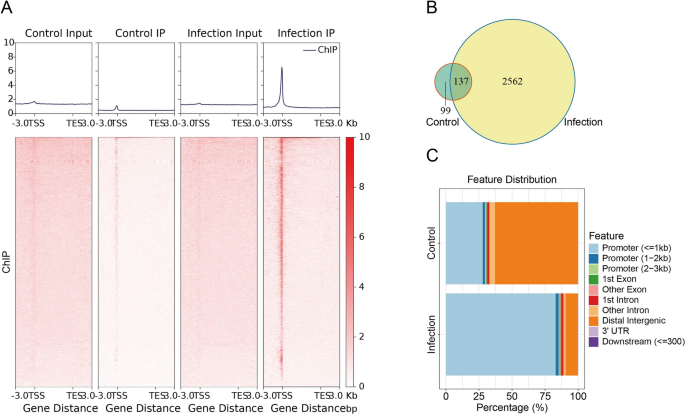
Fig 2.Egr-1结合的靶基因鉴定及生物信息学分析
3. Egr-1结合基因与感染诱导基因的关联分析 ? ?
为了更好地了解Egr-1在细菌性脑膜炎发展中的调节作用,采用生物信息学方法进行了感染组中启动子区富集的独特peak峰的GO功能分析和KEGG富集分析,结果表明,脑膜炎型大肠杆菌诱导的Egr-1具有强烈破坏血脑屏障完整性和增加感染期间炎症反应的能力。结合研究者先前转录组数据发现,人脑微血管内皮细胞(hBMEC)在感染脑膜炎型大肠杆菌后,有366个基因的表达增加了≥2倍或降低了≤0.5倍。通过将这些显著改变的基因与2,322个在启动子区域富集的独特峰相结合,研究者筛选出了111个可能直接受Egr-1调控的候选转录靶点。由于富集于调节细胞骨架和血管生成途径的基因与BBB完整性调节密切相关,研究者提取了调节细胞骨架和血管生成的基因集合,并利用STRING数据库生成了蛋白质-蛋白质相互作用网络。在参与细胞骨架调节的相互作用蛋白质群中,PDGFB显示出最显著的折叠变化,而在参与血管生成调节的相互作用蛋白质群中,ANGPTL4和VEGFA显示出最显著的折叠变化。这些发现表明,PDGFB、ANGPTL4和VEGFA是Egr-1影响BBB通透性的关键靶点(Fig 3)。
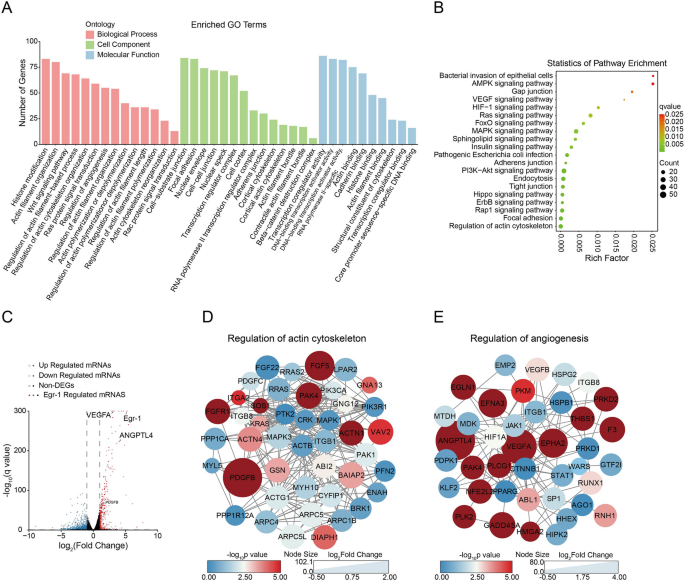
Fig 3.Egr-1靶基因的生物信息学分析
4. 脑膜炎型大肠杆菌诱导的细胞骨架改变需要 Egr-1 激活
在这项研究中,研究者发现Egr-1在感染过程中对BMEC的细胞骨架重排起着关键作用。通过CRISPR/Cas9技术,研究者成功地在hBMEC中基因敲除Egr-1。在野生型细胞中,感染导致Egr-1表达上调,而敲除Egr-1的细胞中未观察到Egr-1表达。在未感染的野生型细胞中,F-actin纤维组装良好且内部拉伸均匀;而在感染的野生型细胞中,F-actin呈分散状态,细胞骨架被破坏和解体,导致细胞变形。然而,在Egr-1被敲除的细胞中,感染并未导致细胞骨架F-actin纤维的明显变化。此外,研究者发现,在感染1小时后,野生型细胞中RhoA、Rac1和Cdc42的GTP结合(活跃)形式增加,但在Egr-1敲除细胞中,这种激活作用得到了很大程度的抑制。这些发现表明,脑膜炎型大肠杆菌诱导的hBMEC细胞骨架改变需要Egr-1的表达(Fig 4)。
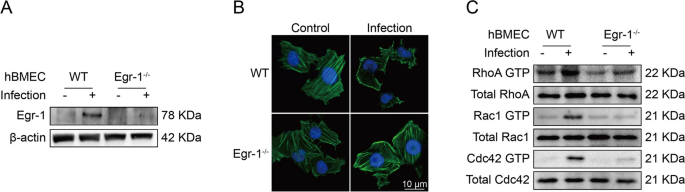
Fig 4. Egr-1激活有助于脑膜炎型大肠杆菌诱导的细胞骨架纤维分解
5. VEGFA、PDGFB和ANGPTL4表达受Egr-1调节
生物信息学分析表明,Egr-1通过其潜在靶基因VEGFA、PDGFB和ANGPTL4调节 actin细胞骨架和血管生成。ChIP-seq数据显示,感染组中VEGFA、PDGFB和ANGPTL4的启动子区域存在显著的Egr-1结合峰,而对照组几乎没有结合峰(Fig 5A)。因此,研究者假设VEGFA、PDGFB和ANGPTL4可能是脑膜炎型大肠杆菌感染hBMEC的Egr-1的潜在靶基因。随后,研究者进行了ChIP-PCR实验,结果显示,细菌处理后的hBMEC中Egr-1与VEGFA、PDGFB和ANGPTL4启动子的结合显著增加(Fig 5B)。此外,RT-PCR和Western blot显示,Egr-1敲除在感染的hBMEC中部分抑制了VEGFA、PDGFB和ANGPTL4的转录和表达水平(Fig 5C和5D)。这些结果共同表明,Egr-1激活对于脑膜大肠杆菌感染的hBMEC中VEGFA、PDGFB和ANGPTL4的上调是必需的。
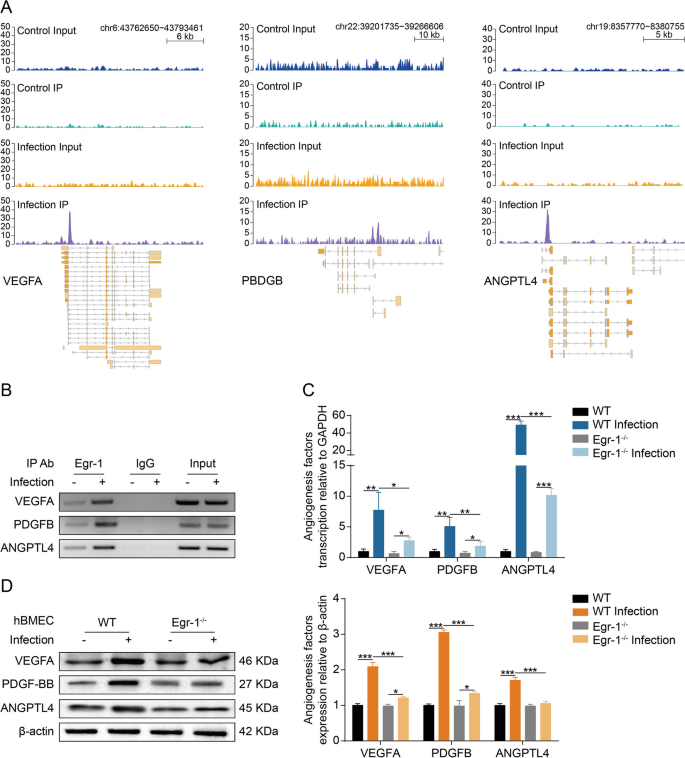
Fig 5.Egr-1是VEGFA、PDGFB和ANGPTL4诱导所必需的
6. Egr-1是破坏TJ蛋白的关键前提条件
脑膜炎型大肠杆菌感染可诱导VEGFA、PDGFB和ANGPTL4表达,从而加重感染导致的血脑屏障(BBB)功能障碍。然而,在Egr-1敲除的hBMEC中,由脑膜炎型大肠杆菌引起的VEGFA、PDGFB和ANGPTL4的诱导作用得到了很大程度的抑制。此外,研究发现Egr-1激活可能影响hBMEC单层的完整性。在Egr-1敲除和野生型hBMEC中,对TJ蛋白(包括ZO-1、紧密连接蛋白和Claudin-5)的调控进行了比较。结果显示,与对照组相比,Egr-1敲除的hBMEC在感染后TJ蛋白的下调在mRNA和蛋白质水平上明显减轻。此外,通过免疫荧光分析发现,Egr-1敲除的hBMEC在感染后TJ蛋白的分布和表达受损程度明显减轻。这些发现表明,脑膜炎型大肠杆菌感染引起的血脑屏障完整性破坏依赖于Egr-1的激活(Fig 6)。
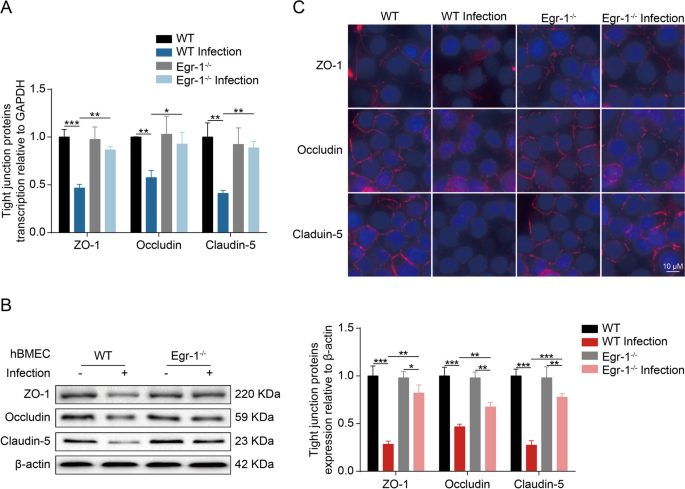
Fig 6.hBMEC中的Egr-1敲除可抑制脑膜炎型大肠杆菌诱导的 TJ 蛋白降解
7. Egr-1 通过增加 VEGFA、PDGFB 和 ANGPTL4 表达来增强 BBB 通透性
在本研究中,研究者发现Egr-1在小鼠脑血管内皮细胞中通过调节VEGFA、PDGFB和ANGPTL4的产生,参与了脑膜炎型大肠杆菌感染引起的血脑屏障破坏。研究者使用Egr-1基因敲除小鼠(Egr-1?/?)来评估Egr-1对VEGFA、PDGFB和ANGPTL4生产的影响。通过酶联免疫吸附试验(ELISA),研究者发现感染导致的VEGFA、PDGFB和ANGPTL4在Egr-1?/?小鼠脑中的上调明显低于野生型(WT)小鼠(Fig 7A)。此外,研究者通过RT-PCR和Western blot检测了小鼠脑中紧密连接蛋白(ZO-1、Occludin和Claudin-5)的表达。在细菌感染的WT小鼠中,这些蛋白的转录和表达水平显著降低,而在Egr-1?/?小鼠中,这种降解过程被显著逆转(Fig 7B和7C)。免疫荧光染色结果显示,感染WT小鼠的血管内皮层分布不连续、不规则或出现间隙,而在感染Egr-1?/?小鼠中,这些紧密连接蛋白的不良影响得到了很好的修复(Fig 7D)。此外,研究者还使用伊文思蓝荧光法评估小鼠血脑屏障的通透性,发现感染Egr-1?/?小鼠的脑部伊文思蓝进入量明显低于感染WT小鼠(Fig 7E)。这些数据表明,Egr-1是大肠杆菌脑膜炎中血脑屏障破坏的重要因素。
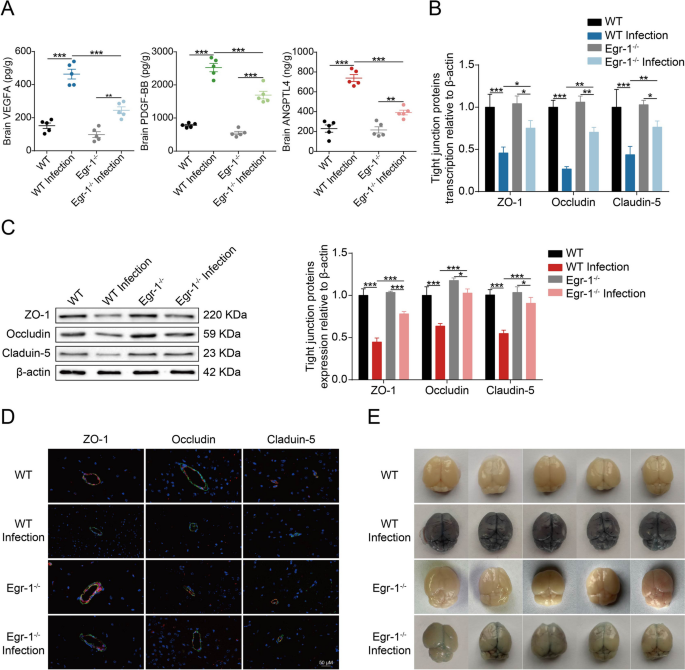
Fig 7.小鼠中Egr-1敲除可抑制脑膜炎型大肠杆菌诱导的BBB破坏
8. Egr-1对脑膜炎型大肠杆菌诱导的神经炎症的作用
为了进一步研究Egr-1对由脑膜炎型大肠杆菌引起的中枢神经系统炎症的影响,研究者使用电化学发光法评估了Egr-1在体内感染过程中调节细胞因子和趋化因子生产的能力。研究者发现,脑部的IL-1β、IL-6、TNF-α、CCL-2和CXCL-2水平在细菌感染的WT小鼠中显著增加;然而,在细菌感染的Egr-1?/?小鼠中,这些细胞因子和趋化因子的上调显著降低(Fig 8A)。白细胞粘附和通过血脑屏障的迁移取决于粘附分子(如ICAM-1和VCAM-1)的激活和表达。因此,研究者分析了细菌感染的WT和Egr-1?/?小鼠脑组织中ICAM-1和VCAM-1的表达。研究者观察到细菌感染的WT小鼠脑部ICAM-1和VCAM-1的表达上调,而在细菌感染的Egr-1?/?小鼠中没有观察到这种上调(Fig 8B)。通过免疫荧光分析了ICAM-1和VCAM-1的表达,与细菌感染的Egr-1?/?小鼠相比,细菌感染的WT小鼠中ICAM-1和VCAM-1的表达增加,并与CD34标记的血管共定位(Fig 8C)。这些证据表明,脑膜炎型大肠杆菌感染诱导的ICAM-1和VCAM-1上调依赖于Egr-1的激活。此外,对对照小鼠和脑膜炎型大肠杆菌感染的WT和Egr-1?/?小鼠进行脑部切片检查。组织病理学检查显示,细菌感染后WT小鼠脑膜明显增厚,伴有相应的炎症细胞聚集,但这种组织学病变在Egr-1?/?小鼠中得到了缓解(Fig 8D)。考虑到Egr-1对脑膜炎型大肠杆菌感染小鼠血脑屏障完整性和神经炎症加重的不利影响,研究者继续研究Egr-1基因敲除是否可以提供保护。实验结果显示,在未感染的WT和Egr-1?/?小鼠中,所有小鼠在观察期内存活。然而,在感染了细菌的野生型小鼠中,死亡率达到了100%。相比之下,Egr-1基因敲除对60%的感染细菌的Egr-1?/?小鼠提供了保护作用(Fig 8E)。这些发现强烈支持Egr-1在脑膜炎型大肠杆菌引起的神经炎症和死亡中的关键作用。
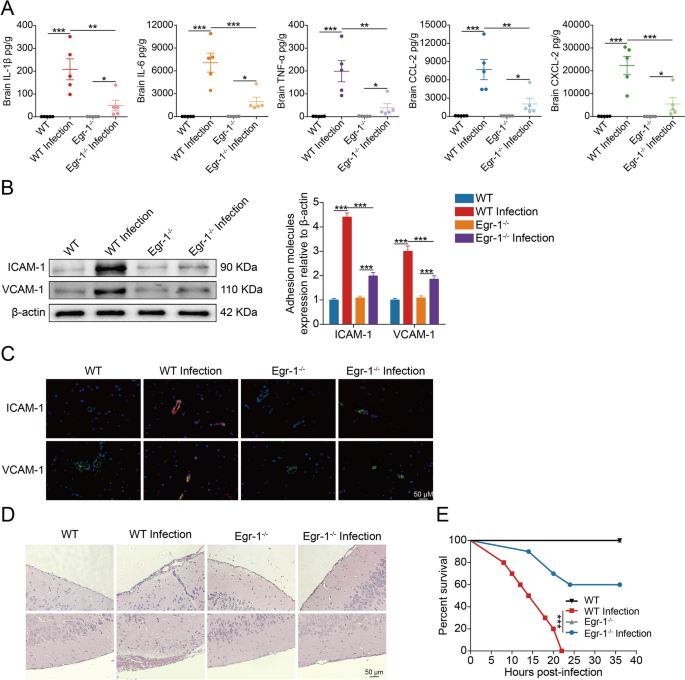
Fig 8.小鼠中Egr-1敲除可减少神经炎症并提高脑膜炎型大肠杆菌感染的存活率
? 总? ?结??
该研究发现,脑膜炎型大肠杆菌能迅速诱导关键宿主转录因子Egr-1的表达。Egr-1通过结合其调节元件,改变细胞骨架和降解紧密连接蛋白,最终导致血脑屏障的破裂。此外,该研究还发现Egr-1是脑膜炎型大肠杆菌感染引发神经炎症的关键调节器。这些结果表明,针对Egr-1的治疗可能代表治疗细菌性脑膜炎的有前途的策略。
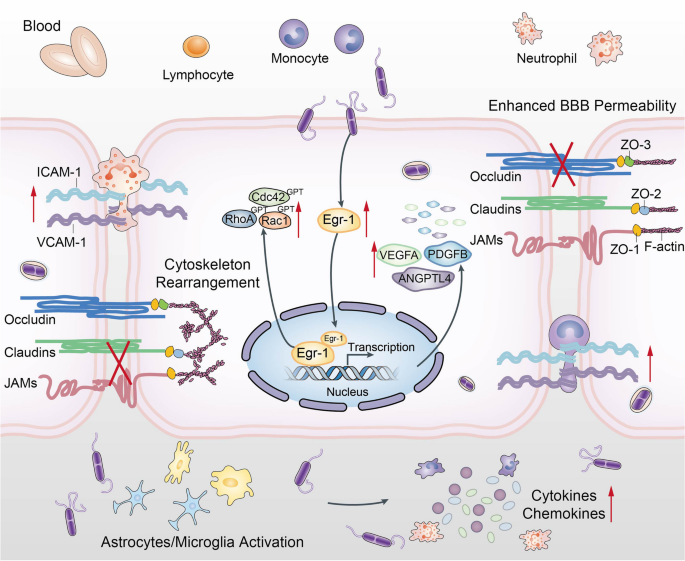
Fig 9.脑膜炎型大肠杆菌诱导Egr-1破坏BBB以及神经炎症反应的示意图
·? 爱基百客王牌产品ChIP-seq简介 ·?
·?ChIP-seq相关介绍?·
ChIP-seq技术将染色质免疫共沉淀和二代测序技术结合,是研究体内蛋白质与DNA相互作用的有力工具,可用于组蛋白修饰、RNA聚合酶、转录因子和辅因子以及G4链体(G4)等方面的研究,技术成熟稳定。爱基百客ChIP-seq可提供:
-
ChIP-seq测序分析
Peak分析:?Peak注释和分布分析,Peak关联基因的GO、KEGG的注释和富集分析, 转录因子和Motif分析等。
多样本差异分析:差异 Peak 分布情况统计,差异 Peak 关联基因GO、KEGG 功能注释与富集,转录因子预测,Motif 预测等。
·??后续验证
01 ChIP-qPCR
分析组蛋白修饰/转录因子与染色质区域的结合情况,揭示染色质状态和基因表达调控之间的关系,真实反映结合特性。
02 EMSA
基于DNA-蛋白质复合体在聚丙烯酰胺凝胶电泳(PAGE)中的迁移率不同,检测活化的与DNA结合的蛋白转录或调节因子。
03 双荧光素酶报告实验
检测转录因子与靶启动子的特异结合。
-
ChIP-seq+转录组关联分析
ChIP-seq和转录组关联分析可以做以下2个方面的研究:
1、DNA结合蛋白和基因表达调控:通过ChIP-seq技术可以确定DNA结合蛋白(如转录因子)的结合位点,然后与转录组数据结合分析,可以获得转录因子直接调控的靶基因,为全面理解转录因子调控功能提供依据。
2、组蛋白修饰和基因表达:ChIP-seq可以用于鉴定组蛋白修饰的位点,结合转录组数据可以了解这些修饰对基因表达的影响。
·??爱基百客ChIP-seq三大优势
优势一:项目经验丰富,研究物种200+种,累计实验2000余次。全面覆盖医口和农口等不同样本,不惧特殊样本(如脂肪组织、高淀粉组织和真菌类),抗体经验也极其丰富(多种组蛋白修饰、转录因子、标签抗体以及p300和RNApol II等均有涉及);
优势二:提供前期实验设计、测序、分析以及后期验证(ChIP-qPCR、EMSA)一站式服务;
优势三:项目文章多次发表于Cancer Cell、Circulation、Nature Plants、Nature Metabolism以及Plant Cell等国际知名期刊。
本文来自互联网用户投稿,该文观点仅代表作者本人,不代表本站立场。本站仅提供信息存储空间服务,不拥有所有权,不承担相关法律责任。 如若内容造成侵权/违法违规/事实不符,请联系我的编程经验分享网邮箱:chenni525@qq.com进行投诉反馈,一经查实,立即删除!